
Author: Eric L. Mills
Department of Oceanography, Dalhousie University (Halifax, Nova Scotia, Canada)
History of Science and Technology Programme, University of King’s College (Halifax, Nova Scotia, Canada)
Abstract
After the publication of the last Challenger Report in 1895, deep-sea biology fell out of favor with marine biologists. It was replaced by embryology and physiology in the mainline biological sciences and by attention to biological oceanography, especially the marine production cycle, within oceanography. Only after World War Two did deep-sea biology come back into favor, beginning with the round-the-world cruises of the Swedish Albatross (1947-1948) and the Danish Galathea (1950-1952). In the United States, one renewal of deep-sea biology occurred, rather unexpectedly, at Woods Hole Oceanographic Institution in the 1960s, not long after the first International Oceanographic Congress of 1959. How this came about proves to be a mix of opportunism, chance, and Realpolitik that turned a natural history study into an ecological science.
Introduction: resurrecting deep-sea biology
Deep-sea biology captured scientific imaginations for close to sixty years in the nineteenth century, promising to throw light on the origins of biological diversity, to give evidence of “living fossils”, and to reveal adaptations to life under great pressures, lack of light, and especially lack of food. Several great expeditions to consider these scientific problems, always linked to scientific and political ambitions, took place after the mid-nineteenth century, the greatest of them the Challenger Expedition (1872-1876). And yet, after 1900 deep-sea biology disappeared from the scientific stage and the lights dimmed on biological work in deep water for close to half a century. After World War Two, a partial rejuvenation of deep-sea biology took place in the hands of Scandinavian marine scientists, then in the 1950s and early 1960s the field fully emerged from the shadows and took its place among the rapidly expanding ecological sciences.
I was a witness to some of the changes that took place in the 1960s, and later, in a small way, a participant in the new science. In the paper to follow, I attempt to give a personal view of how the changes came about, but in a larger context, one that I certainly did not understand when the study of deep-sea diversity began to unfold in the early 1960s.
Background of work in the deep sea
The Challenger Expedition of 1872-1876 gave firm evidence that the ocean was full of life to the greatest depths sampled, in this case 3125 fathoms (about 5176 meters).[1] For a time, the example set by this great expedition stimulated more work by Germans, the French, and especially Americans. In the Americas, Alexander Agassiz (1835-1910) was especially influential, because of his training as a mining engineer and his wealth, in carrying on deep-sea research.[2] Using the U.S. Coast Survey steamer Blake from 1877 into the 1880s,[3] then the new U.S. Fish Commission steamer Albatross from 1891-1900 and in 1904-1905, he investigated the deep-sea benthos and mid-water plankton using the most sophisticated technology available in his time.[4] In large part, Agassiz’s results were extensions of those from Challenger, but he was highly influential in introducing new techniques (such as wire rope for trawling – borrowed from mining technology) and, because he used his own wealth, in providing an example of the private patronage of science.
A contemporary of Agassiz, Albert 1er, Prince of Monaco (1848-1922), also wealthy and influential, carried out extensive work on the deep ocean from his yachts, especially the magnificent Princesse-Alice II.[5] Using a variety of techniques, ranging from giant trawls to baited traps, the Challenger results, once again, were extended to new areas and new species. The Prince of Monaco’s successful trawling operation at 6035 meters in the Atlantic south of the Cape Verde Islands in August 1901, exceeding Challenger’s deepest station, was the deepest sampling of the abyssal depths until 1947.
Agassiz’s work, along with that of the Prince of Monaco and a few others, was exceptional and less and less characteristic of the main directions taken by biology between the 1880s and the Second World War. We find the bulk of publications in the biological marine sciences during those years in physiology, embryology, plankton dynamics, and fisheries science. Biology, professionalizing in universities and in government, took direction on the one hand toward cellular structure and mechanism and on another toward solving practical problems of fisheries. Deep-sea biology became a side issue, pursued by very few.
Deep-sea biology revives
When deep-sea biology reappeared after the Second World War it did so not out of biology but out of radiochemistry and sedimentology. The Swedish radiochemist Hans Pettersson (1888-1966), son of the influential Otto Pettersson, a founder of the International Council for the Exploration of the Sea, had wanted to mount a Swedish deep-sea expedition since the 1930s, one devoted to the sediments.[6] Subsidized by the Swedish shipping firm Bröstrom, the training schooner Albatross (Figure 1) was prepared and provisioned, with the aim of working mainly on deep-sea sediments using the newly-developed techniques of piston-coring and seismic profiling.[7]

There was little biological work during the circumnavigation by Albatross that took place between 4 July 1947 and 3 October 1948 (Figure 2) until the last three months when the marine biologist Orvar Nybelin supervised deep trawling to more than 7000 meters in the Puerto Rican Trench, recovering fish and invertebrates (including worms and holothurians) from the greatest depths yet sampled.[8]

There was also pre-war interest in deep-sea investigation in Denmark. It had to be delayed until five years after the war, when an expedition fund was set up, equipment was transferred from Albatross, and a ship was bought, a retired British Royal Navy sloop named H.M.S. Leith, which was renamed Galathea (Figure 3) after its nineteenth century predecessor.[9]

The instigator of what became the Galathea Expedition was the Danish zoologist Anton Bruun (1901-1961), who had circumnavigated the world between 1928 and 1930 on the Danish Dana II, on which the studies had been mainly of fish and plankton, and had studied near-shore benthos from the yacht Atlantide off the West African coast in 1945-1946. Bruun had wide-ranging interests in the sea, including the possible existence of sea-serpents, and his enthusiasm captured the interest of the Danish public through the journalist Haakon Mielche, resulting in widespread support for another Danish circumnavigation, this one devoted to the deep ocean.[10]
The Galathea Expedition took place from 15 October 1950 to 29 June 1952 (Figure 4), devoted to the study of trench faunas using trawling and quantitative studies of the abyssal fauna using grab samplers; there were also studies of plankton (including early use of C-14 to estimate production), hydrography, magnetics, terrestrial biology and anthropology – making it close to a twentieth century analogue of the Challenger Expedition.
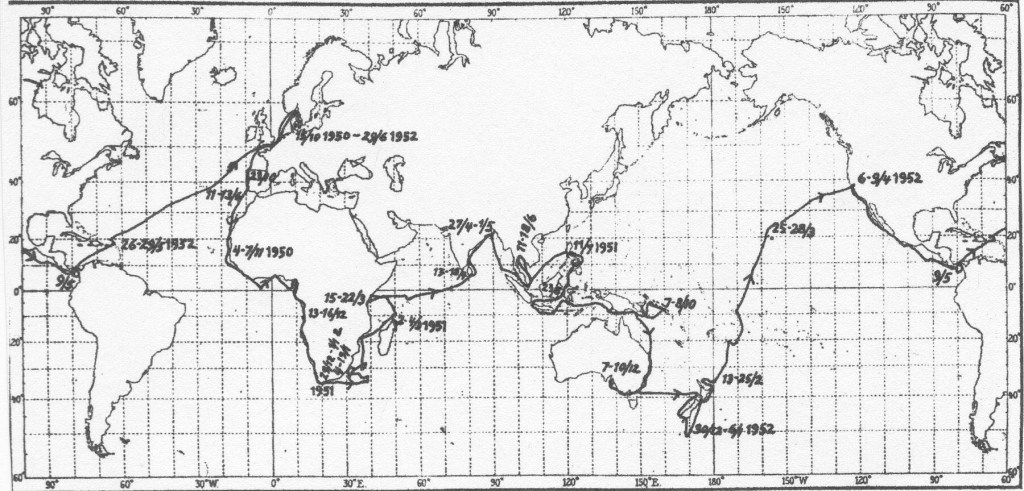
The Galathea biologists trawled successfully at 10,190 meters in the Philippine Trench – a noteworthy technical accomplishment – and in four more trenches before the cruise ended, finding more than 115 species at hadal depths (below 6000 meters), defined a hadal fauna,[11] and provided early estimates (from 65 grab samples at abyssal depths) of the decreasing abundance of animals as depth increased. They also discovered the primitive monoplacophoran mollusc Neopilina at 3600 meters just north of the Gulf of Panama, providing evidence that at least one representative of an ancient group still existed in the deep sea.
Nearly simultaneously, marine biologists from the Soviet Union began work at abyssal and hadal depths, at first largely in the Pacific. The ship Vitiaz was sent from European Russia to the Russian Far Eastern seas in 1949 (Figures 5, 6).
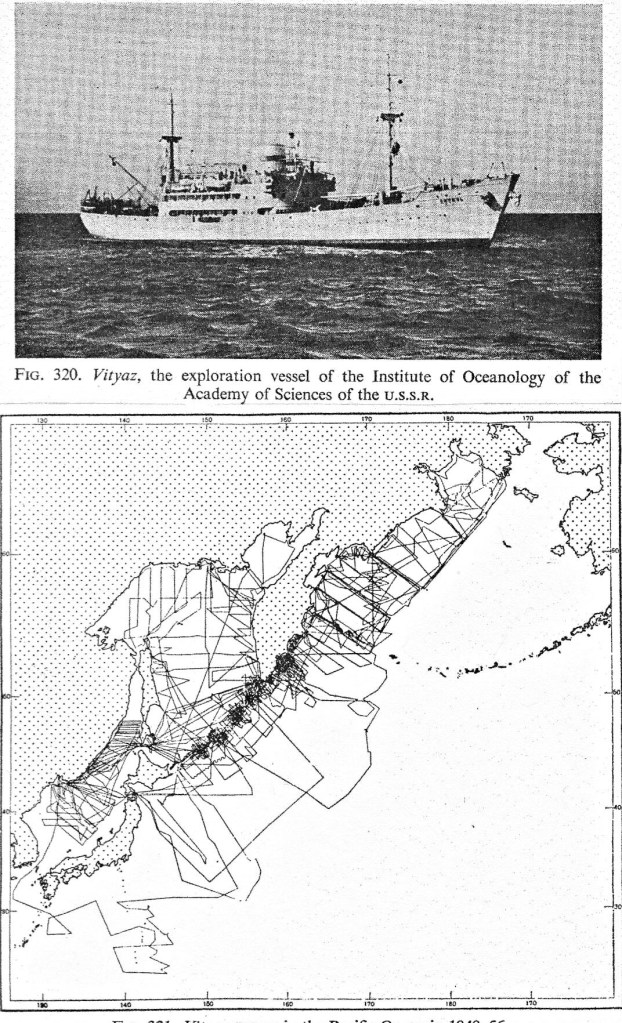
There it became involved in an extensive program of grab sampling throughout the area (including the Kurile-Kamchatka Trench) from then until 1956. After 1956, Vitiaz was moved farther afield, working as far east in the Pacific as North America and as far south as New Zealand during the International Geophysical Year (IGY) of 1957-59 and through 1962. It is now a museum ship in Kaliningrad (Figure 6).

During this time, the Russian research vessels Ob and Mikhail Lomonosov added information from other oceans.[12] Meanwhile, in the United States, Robert J. Menzies (1923-1976) began a long series of studies of the systematics and distribution of deep-sea animals, concentrating mainly on isopods and on the primitive mollusc Neopilina, working intensively primarily in the Pacific and also the Antarctic from the mid 1950s through the early 1970s.
The Russian work in the deep ocean was aimed at the “biological structure of the ocean” according to the instigator of much of this work, the Academician Lev Zenkevitch.[13] Systematic, thorough, and geographically wide-spread, the Russian work in the Pacific gave the first extensive information on benthic abundance over large areas, along with a large volume of results on the taxonomy and zoogeography of the animals collected in the deep sea (Figure 7).

Taken together with the results of the Swedish and Danish investigations, the belief grew that there was, as expected, a striking decrease in biomass, abundance, and species diversity as depth increased from near shore to abyssal and hadal depths. But shortly after the end of the IGY in 1959, these results were overthrown by a study of the abyssal fauna along a transect in the North Atlantic Ocean.
The Massachusetts – Bermuda transect
After some preliminary trials, a totally new study of the deep-sea fauna began on a cruise of Woods Hole Oceanographic Institution’s (WHOI) research vessel Atlantis (Figure 8) from 20-25 May 1961 along a transect across the North American Basin from Gay Head on Martha’s Vineyard, Massachusetts, to Bermuda.[14]

The investigators were Howard L. Sanders (1921-2001) (Figure 9) and Robert R. Hessler (1932-2020) (Figure 10) of WHOI, using gear modified and scaled up from inshore versions, an anchor dredge for animal abundances (Figure 11), and later an epibenthic sled (Figure 12) to give larger numbers for species diversity studies.





Figure 9. Howard Sanders at work on the deck of Atlantis II, August 1966. Photograph by Eric Mills; Figure 10. Robert R. Hessler (center) in Tahiti during Scripps Institution of Oceanography’s Operation South Tow, January 1972. Photograph by Eric Mills; Figure 11. An anchor dredge, used for quantitative assessments of abyssal animals, typical of those used by Sanders and Hessler along the Gay Head-Bermuda transect. Photograph by Eric Mills; Figure 12. An epibenthic sled typical of those used by Sanders and Hessler along the Gay Head-Bermuda transect. The original design was by Robert Hessler with contributions by George Hampson and Rudolf Scheltema. Use of sleds revealed the very high diversity of the deep-sea benthic fauna. Robert Hessler is at extreme right and George Hampson second from right. Photograph by Eric Mills; Figure 13. Processing a benthic sample on Atlantis in October 1961. George Hampson is on the left, Howard Sanders on the right. Photograph by Eric Mills.
There were new processing devices too (Figure 13), including a calibrated garbage can enabling the volume of mud (and thus the surface area covered in sampling by the anchor dredge) to be measured, and unusually fine screens with mesh size 0.42 mm (most previous sampling had used 1 mm or larger screens, although Russian investigators on Vitiaz in 1954-55 had begun to use 0.5 mm netting to separate what they called the “microfauna” from the sediment).
These simple but very effective sampling and processing techniques rapidly revealed unexpected abundances of animals in the deep-sea,[15] and shortly thereafter, when taxonomic work began, unexpectedly high species diversity, as high in some places as in the richest terrestrial environments, tropical forests. These striking results were borne out by a succession of cruises during the next decade along the Massachusetts – Bermuda transect and then into other oceans, allowing Sanders to document abundances and diversity widely, to develop ways of expressing and comparing species diversity measures, and to promote a hypothesis to explain high diversity in the deep sea. Sanders’ stability-time hypothesis postulated that the stable conditions of the deep sea and the age of the abyssal environment enabled competition and niche differentiation to go to completion, resulting in the presence of high numbers of species under seemingly uniform conditions.[16] These studies came at a time of great interest by ecologists in niche relations and the development and maintenance of diversity, and thus fitted neatly into the preoccupations of contemporary theoretical ecologists.

Although the stability-time hypothesis was soon contested,[17] the overall results of Sanders’ and Hessler’s work led to a burst of new activity on deep-sea biology, in the hands of a few at first, including Andrew G. Carey at Oregon State University (Figure 14), Fred Grasslé at WHOI and Rutgers, Hessler himself after moving to the Scripps Institution of Oceanography in 1969, then many more researchers right up to the present day.
The beginning of the Sanders / Hessler era
In 1960, Howard Sanders (1921-2001) was best known as the discoverer of the first cephalocarid crustacean, Hutchinsoniella macracantha, a potential modern representative of the ancestral groups giving rise to all of the Crustacea that had been discovered in the mud of Long Island Sound during his doctoral research at Yale from 1951 to 1955.[18]

He was hired full time at WHOI in 1955 under Bostwick (Buck) H. Ketchum (1912-1982) (Figure 15), then senior oceanographer, to expand biological studies at the Institution into benthic ecology, specifically the near-shore and intertidal benthos of the Cape Cod area . At the same time Sanders began to enlarge his work on Hutchinsoniella to include its ecology and the functional morphology of locomotion and feeding.
In the summer of 1959, when Sanders was about to begin work on animal-sediment relations on the flats of Barnstable Harbor, Massachusetts, I was hired to work for him for a few months as a technician. We spent many days in the field collecting quantitative samples of sediment and animals on the Barnstable flats (Figure 16), and many more days processing the samples.

Out of this came a publication showing that many of the apparent carnivores were in fact omnivores, selectively ingesting sediment in various particle ranges along with animal prey items in some cases; that the sediments contained abundant unicellular plants to serve as food; and documenting the life histories of the dominant species.[19] That study was entirely typical of his work at the time, and Sanders seemed poised to go on in the same vein, working on Hutchinsoniella and animal-sediment relations, for some time.
In 1960, Howard Sanders was joined at WHOI by Robert Hessler, fresh from a Ph.D. in paleobiology at the University of Chicago. Hessler’s expertise in trilobite structure and function would enable them to collaborate on functional morphology studies of Hutchinsoniella, with the aim of throwing more light on trilobite-crustacean relationships and the phylogenetic position of cephalocarids. But although the work with cephalocarids did take place, within a few months Sanders and Hessler were at sea between Massachusetts and Bermuda beginning the deep-sea work for which both became famous, leaving behind much of the inshore benthic work. What caused this sudden change of direction?
Innovation at Woods Hole Oceanographic Institution
Late in the summer of 1959, just before he left Woods Hole to attend the first International Oceanographic Congress in New York, Howard Sanders told me that he was being put under pressure to move into deep-sea biology. He made it clear that this had not been his decision, and that he wasn’t happy about it. But move he did, and within two years his new research area was giving unexpected results, leading to revolutionary changes in deep-sea biology. Sanders soon had reason to be grateful for this forced change of direction.
Many years later, in 1980 at a reception in Woods Hole, I met Buck Ketchum, then Associate Director of WHOI. Somehow my first experiences in Woods Hole in 1959 and what Howard Sanders had told me then about his change of research direction came up in the conversation. When I mentioned what Sanders had said, Ketchum told me that it had been he who gave the instructions to move into deep-sea biology. I left it at that, leaving the question “Why?” unasked, but the memory remained with me.
New insight into these events came with my attempt to unravel the “resurrection” (perhaps better the “revival”) of deep-sea biology in the 1960s, and was stimulated particularly by reading Hamblin’s book Poison in the Well[20] about disposal of nuclear wastes in the sea, and WHOI directors’ reports for the late 1950s and 1960s.
In the mid-1950s, oceanic plankton production research under the direction of Buck Ketchum at WHOI began to expand seaward. By 1957 it had moved onto the continental shelf and beyond, from its start in coastal waters.[21] In 1958, Ketchum got funding from the U.S. Atomic Energy Commission to extend the work across the Sargasso Sea to Bermuda. This took place in the broad context of debate in the United States and elsewhere about the utility and safety of dumping nuclear wastes in the deep sea, a debate in which Ketchum played a part on the domestic and international stages. Although by 1960/1961 the U.S. Atomic Energy Commission had given up ocean dumping and decided to concentrate on land disposal, many marine scientists had, or were attempting to get, research funding from the AEC.[22]
Ketchum’s role in these events, as a radio-ecologist, plant physiologist, and spokesman for marine science, was significant. For example, in 1955, well before ocean dumping ended, he and his WHOI colleague Allyn Vine attempted to interest the U.S. AEC in ocean circulation and marine ecology, making the case that very little was known about the distribution and fate of radioisotopes in marine organisms once they began to spread in the sea after dumping or nuclear explosions.[23] That year Ketchum, as a specialist in marine ecology, spoke in Geneva at the First International Congress on Peaceful Uses of Atomic Energy. In 1956 he attended the Conference on Biological Effects of Atomic Radiation in Princeton, NJ, again speaking on the biological effects of atomic radiation on marine organisms. A year later, in 1957, he was in Göteborg, Sweden, at a meeting of the United Nations Special Committee on Effects of Atomic Radiation, speaking on the concentration of radioisotopes by marine organisms.[24]
By 1959, Ketchum and the chemist Vaughan Bowen of WHOI were getting research funding from the AEC’s Division of Biology and Medicine for a program on the distribution of isotopes in the sea, aimed at determining the effects of currents, mixing, biological concentration of the isotopes, and the effects of plankton vertical migration on radioisotopes in marine organisms.[25] Ketchum was also a member of the working group on artificial radioactivity in the sea that was established under the National Academy of Sciences Committee on Oceanography (NASCO) under the chairmanship of Harrison Brown that reported in 1959, recommending ecosystem studies in deep water and, overall, an increase in research on deep-sea biology.[26] That year, 1959, in the WHOI director’s report, Ketchum commented, in his report on marine biology at WHOI, that “the studies of the bottom communities have been largely confined to the inshore, shallow areas”, hinting at a shift of emphasis in the Department’s work.[27] Within a few months of that statement, perhaps even before it, Howard Sanders had been redirected into deep-sea biology.
Resurrecting deep-sea biology
There were at least four factors at work in the redirection of Howard Sanders’ work from inshore benthic ecology to deep-sea biology and the conversion of Challenger-style natural history of the deep sea into twentieth century abyssal ecology. The first was certainly Ketchum’s involvement internationally and within the United States with the issue of the disposal of radioactive wastes, and more generally with the overall fate of radionuclides in the sea, especially in organisms and food-chain pathways. At the same time, WHOI’s biology program (under Ketchum) was moving offshore toward Bermuda in the late 1950s, but that program lacked information on deep-sea benthic animals. Initially, a driving force here may have been the possibility of U.S. Atomic Energy Commission funding, but it is also clear that many WHOI research programs were being redirected toward the open sea with or without that funding and that Ketchum’s (in which Sanders was to play a part ) was one of these. By 1959 planning for a replacement of the old and worn-out Atlantis was well under way and a grant had been received from the National Science Foundation for that purpose. The new ship, Atlantis II, which came into service in 1963 (Figure 17), gave the possibility of reliable work on the global oceans and of being able to enlarge WHOI’s research programs in a way not possible with the older ship. It was a part of the expansion of work at WHOI into which Ketchum’s biology program fitted in an important way.

Finally, there are Sanders’ own background and interests. It is not surprising that his early research at WHOI grew out of his work at Yale on the near-shore benthic ecology of Long Island Sound, centered on feeding ecology and animal-sediment relationships. But partly because of his discovery of the cephalocarid Hutchinsoniella, Sanders had a close relationship with G. Evelyn Hutchinson (1903-1991), the most influential ecologist of his generation. Hutchinson and his students focused their work on niche relationships and the problem of species diversity in new and very influential ways.[28] Once the high diversity of the deep-sea fauna was revealed by Sanders’ early work, it was natural for him to place the results in the theoretical context that Hutchinson had established and that was pervading ecology in the 1960s.
In closing, there is a historiographic point that can be made too. Using standard sources, it might have been easy to see Howard Sanders’ move into deep-sea biology as a natural evolution of his work on shallow-water benthic ecology. Historical analysis from secondary sources could lead to that conclusion. But the sheer coincidences of my being in his laboratory one August afternoon in 1959, of hearing his dismay at being redirected away from shallow water studies, and of bringing up that comment with Buck Ketchum 21 years later, gives us a different story, one that standard sources would not have revealed. The resurrection of deep-sea biology in Woods Hole had a human dimension that would be hard to reconstruct without access to the living dramatis personae.
Acknowledgements
I am grateful to my friend and colleague Bob Hessler for reviewing the first version of this paper and for providing new information on his work with Howard Sanders. His loss in October 2020 severed one of the last remaining links to the Sanders-Hessler era in deep-sea benthic studies.
Footnotes
[1] For a reinterpretation of the results, see Eric L. Mills, “T.RR. Stebbing, the Challenger and knowledge of deep-sea Amphipoda” in Challenger Expedition Centenary, Edinburgh, September 12 to 20, 1972. Proceedings of the Royal Society of Edinburgh B, 72 (1972): 69-87
[2] Eric L. Mills, “Problems of deep-sea biology: an historical perspective” in The Sea. Volume 8. Deep-Sea Biology, ed. G.T. Rowe (New York: John Wiley and Sons, 1983), 32-46.
[3] Alexander Agassiz, Three Cruises of the U.S. Coast and Geodetic Survey Steamer Blake in the Gulf of Mexico, in the Caribbean Sea, and along the Atlantic Coast of the United States, from 1877to 1880 (London: Sampson Low, 1888), 2 vols.
[4] Eric L. Mills, “Alexander Agassiz, Carl Chun, and the problem of the intermediate fauna” in Oceanography: the Past, eds. M. Sears and D. Merriman (New York: Springer-Verlag, 1980), 360-372.
[5] Mills 1983, 48-58; Eric L. Mills, The Fluid Envelope of our Planet: How the Study of Ocean Currents Became a Science (Toronto: University of Toronto Press, 2009), 164-171.
[6] Hans Pettersson, Westward Ho with the Albatross (London: Macmillan); Pettersson, “The voyage,” Reports of the Swedish Deep-Sea Expedition, 1947-1948 1 (1966): 1-123.
[7] E.T. Christiansson, “How the Albatross became a research vessel,” Reports of the Swedish Deep-Sea Expedition, 1947-1948, 1: 125-142.
[8] Mills 1983, 63-64.
[9] Torben Wolff, Danish Expeditions on the Seven Seas (Copenhagen: Rhodos, 1967); Wolff, “The circumnavigation of the globe by the corvette “Galathea.” An expedition with political, economic and scientific goals” in The Golden Age in Denmark. Art and Culture 1800-1850 (transl. Barbara Haveland), ed. B. Scavenius (Copenhagen: Gyldenhal, 1994), 156-163.
[10] Anton Bruun, Sven Greve, Haakon Mielche and Ragnar Spärck, The Galathea Deep-Sea Expedition 1950-1952 (London: George Allen & Unwin, 1956); Mielche, Round the World with Galathea (London: William Hodge, 1953); Mills 1983, 64-65; Wolff 1967, 251-305.
[11] G.M. Belyaev, Hadal Bottom Fauna of the World Ocean (Jerusalem: Israel Program for Scientific Translation, 1972)
[12] N.G. Vinogradova, “Some problems of the study of deep-sea bottom fauna,” Journal of the Oceanographical Society of Japan, 20th Anniversary Volume 1962: 724-725; Belyaev, v, 1-15.
[13] Vinogradova, 725; L.A. Zenkevitch, Biology of the Seas of the USSR (London & Toronto: George Allen & Unwin, 1963).
[14] H.L. Sanders, R.R. Hessler and G.R. Hampson, “An introduction to the study of deep-sea benthic faunal assemblages along the Gay Head-Bermuda transect,” Deep-Sea Research 12 (1965): 845-867.
[15] Sanders , Hessler & Hampson, 856-862.
[16] R.R. Hessler and H.L. Sanders, “Faunal diversity in the deep sea,” Deep-Sea Research 14 (1967): 65-68; H.L. Sanders, “Marine benthic diversity: a comparative study,” American Naturalist 102 (1968): 243-282; H.L. Sanders, “Benthic marine diversity and the stability-time hypothesis” in Diversity and Stability in Ecological Systems, Brookhaven Symposia in Biology 22 (1969): 71-81; H.L. Sanders and R.R. Hessler, “Ecology of the deep-sea benthos,” Science 163 (1969): 1419-1424.
[17] For a discussion of the controversies and competing hypotheses, see J.D. Gage and P.A. Tyler, Deep-Sea Biology. A Natural History of Organisms at the Deep-Sea Floor (Cambridge: Cambridge University Press, 1999), 201-228; and M.A. Rex and R.J. Etter, Deep-Sea Biodiversity: Pattern and Scale (Cambridge, MA: Harvard University Press, 2010), 60.
[18] H.L. Sanders, “The Cephalocarida, a subclass of Crustacea from Long Island Sound,”Proceedings of the National Academy of Sciences (USA) 41 (1955): 61-66; H.L. Sanders, “The Cephalocarida and crustacean phylogeny,” Systematic Zoology 6 (1959): 112-128.
[19] H.L. Sanders, E.M. Goudsmit, E.L. Mills and G.R. Hampson, “A study of the intertidal fauna of Barnstable Harbor, Massachusetts,” Limnology and Oceanography 7,1 (1962): 63-79.
[20] Jacob D. Hamblin, Poison in the Well: Radioactive Wastes in the Oceans at the Dawn of the Nuclear Age (Rutgers, NJ: Rutgers University Press, 2008).
[21] WHOI, The Woods Hole Oceanographic Institution. Report for the Year 1957 (Woods Hole, MA: Woods Hole Oceanographic Institution, 1958), 20.
[22] WHOI, The Woods Hole Oceanographic Institution. Report for the Year 1956 (Woods Hole, MA: Woods Hole Oceanographic Institution, 1957), 16-17. See especially J.D. Hamblin, “Environmental diplomacy in the Cold War: the disposal of radioactive waste at sea during the 1960s,” International History Review 24, 2 (2002): 348-375; J.D. Hamblin, “Hallowed lords of the sea: scientific authority and radioactive waste in the United States, Britain, and France,” Osiris 21 (2006): 209-228; and Hamblin 2008.
[23] Hamblin 2008, 77-78.
[24] Hamblin 2008, 88-91, 112.
[25] WHOI, The Woods Hole Oceanographic Institution. Report for the Year 1958 (Woods Hole, MA: Woods Hole Oceanographic Institution, 1959), 30; WHOI, The Woods Hole Oceanographic Institution. Report for the Year 1959 (Woods Hole, MA: Woods Hole Oceanographic Institution, 1960), 33; Hamblin 2008, 140.
[26] National Research Council (U.S.). Committee on Oceanography, Oceanography 1960-1970 (Washington, DC: National Academy of Sciences, National Research Council, 1959-1962). See also B.H. Ketchum, “Oceanographic research required in support of radioactive waste disposal” in Radioactive Wastes (Vienna: International Atomic Energy Agency, 1960), 283-291.
[27] WHOI, The Woods Hole Oceanographic Institution. Report for the Year 1959 Woods Hole, MA: Woods Hole Oceanographic Institution, 1960), 37.
[28] For the flavor of Hutchinson’s approach, based on his lectures at Yale, see G.E. Hutchinson, An Introduction to Population Ecology (New Haven, CT: Yale University Press, 1978). On his widespread influence, see Nancy G.Slack, G. Evelyn Hutchinson and the Invention of Modern Ecology”(New Haven, CT: Yale University Press, 2010)

[…] History of Oceanography blog has a new article, “The Abyss: Resurrecting Deep-Sea Biology in the Mid Twentieth Century”. This adds some interesting historical context to Lovecraft’s ‘turn to deeps’ in […]